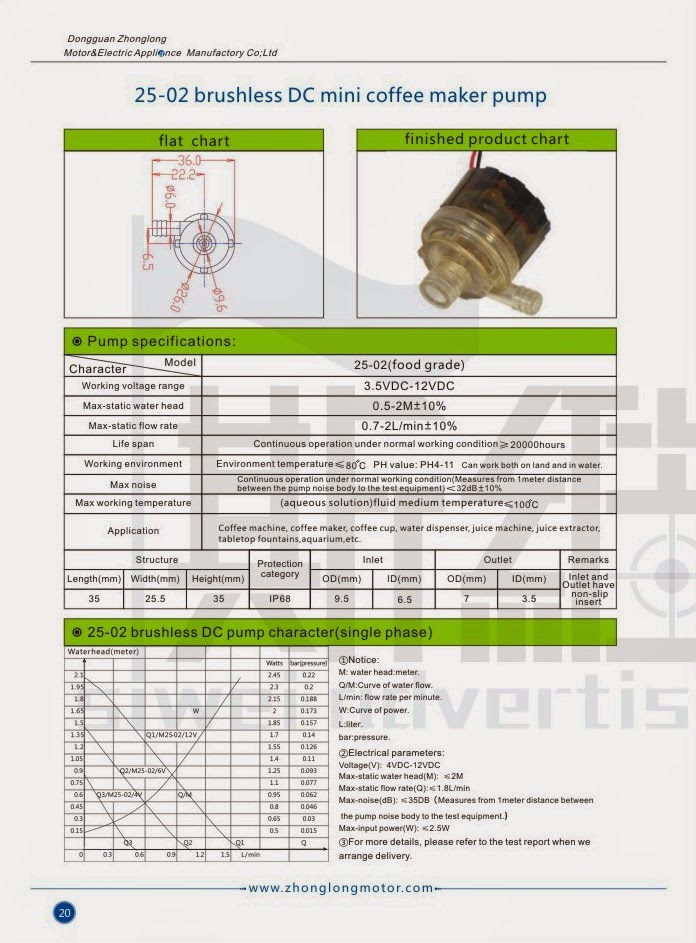
In this article I have explained a protection circuit which may be used for preventing a “dry run” situation in mini coffee dispenser motor pumps, by sensing the slight difference in its wet and dry current consumption levels. The idea was requested by Mr. Ken Adler. Technical Specifications I read with great interest your post titled, […]
Automation Projects
SMS Based Pump Controller with Automatic Dry Run Shut Off
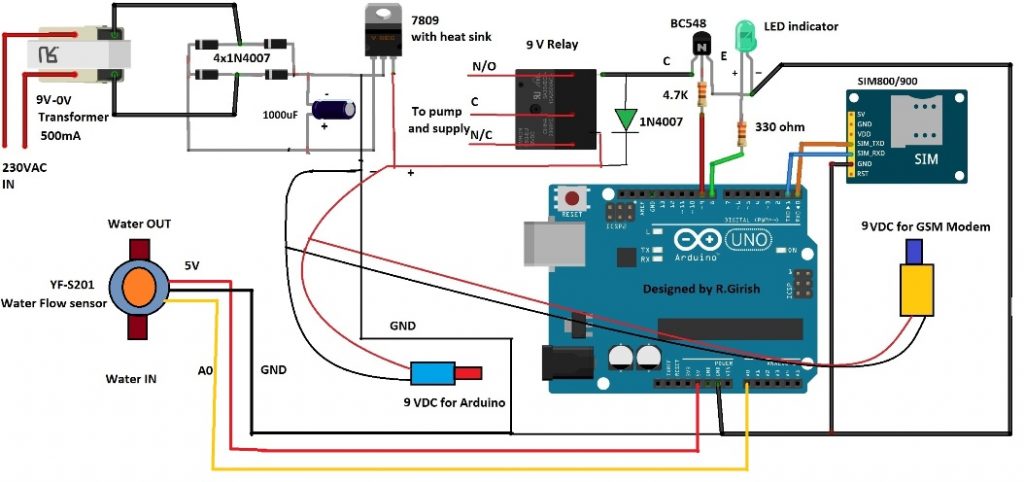
In this post I will show how to construct a SMS based water pump controller with automatic shutdown of pump when no water flow through the pump is detected. We will also construct another much simpler automatic dry run preventer circuit without GSM in the next half of the article. What is Dry Run in […]
Line Follower Robot Circuit using Arduino

In this post I have explained how to build a line follower robot circuit using Arduino, which will run over a specifically drawn line layout and will faithfully follow it as long as it is available and traceable by its sensors. By Navneet Sajwan What is a Line Follower Robot An autonomous robot is a machine […]
Automatic Irrigation Circuit using Arduino

In this post I will show how to construct an automated water irrigation system for small garden using arduino and soil moisture sensor. Introduction The proposed system can monitor the soil moisture level and when soil moisture goes below preset value, the 12V DC pump will be triggered for predetermined period of time. The status […]
7 Automatic Street Light Circuits [Using Relays and Solar Panel]
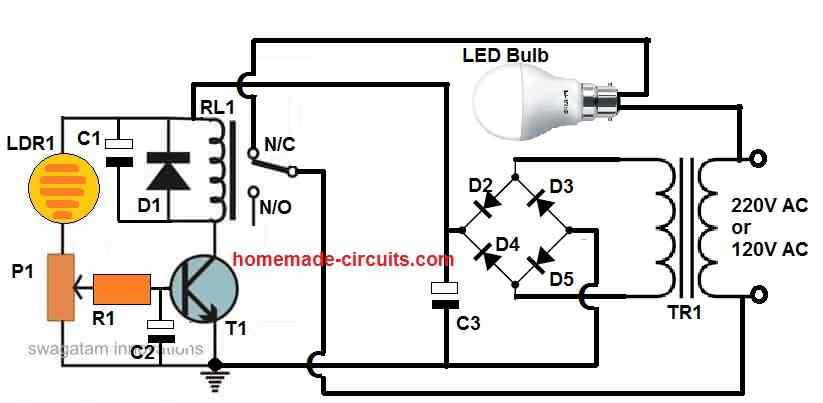
In this article I will elucidate 7 useful yet simple automatic street light circuits using 220 V relays and solar panel. All the presented circuits can be used for automatically switching a lamp ON during night time and OFF during day time. What is an Automatic Street Light System An automatic street light system is […]
Smart Street Lamp Circuit with Motion Sensor

This simple yet highly sophisticated, smart LED street light circuit automatically switches ON a street lamp from low brightness to high brightness whenever it detects a moving object underneath it. This action is initiated only during nighttime, while during daytime the street lamp remains completely shut off. What this Circuit will do This advanced automatic […]